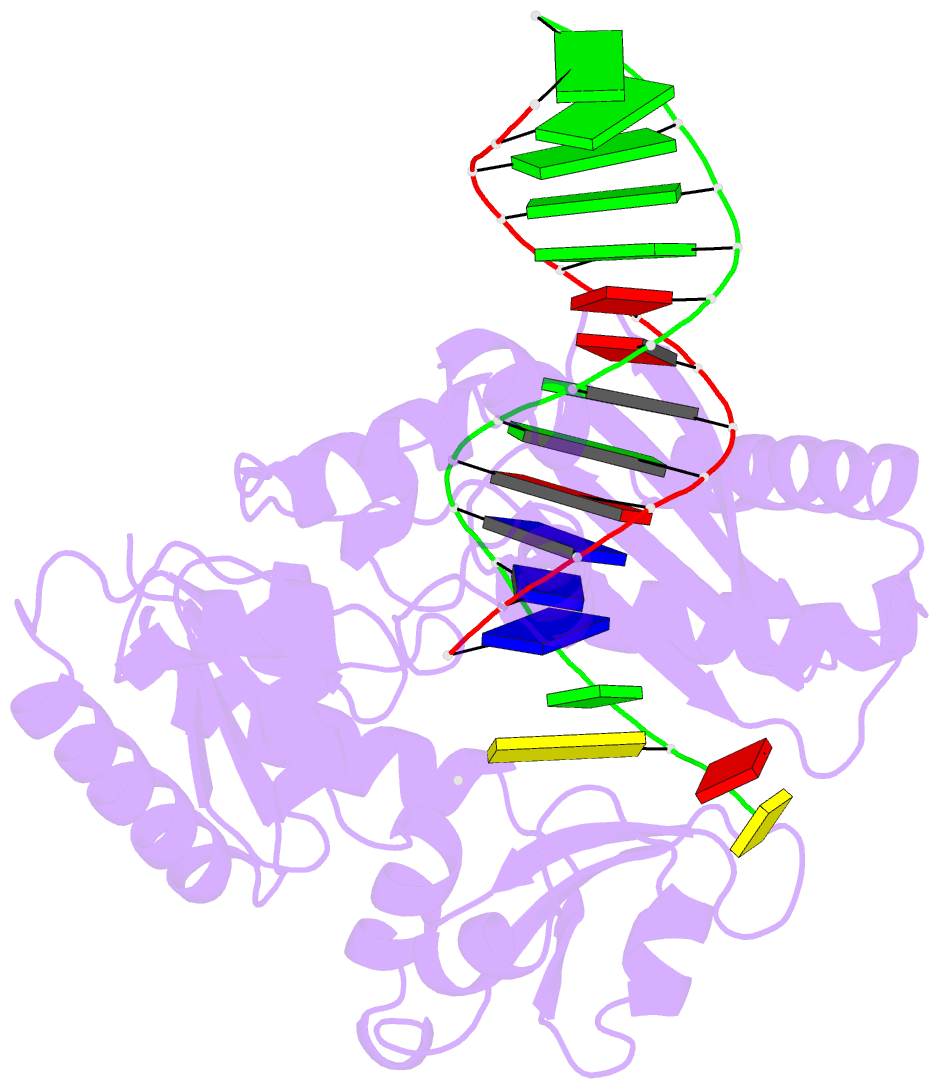
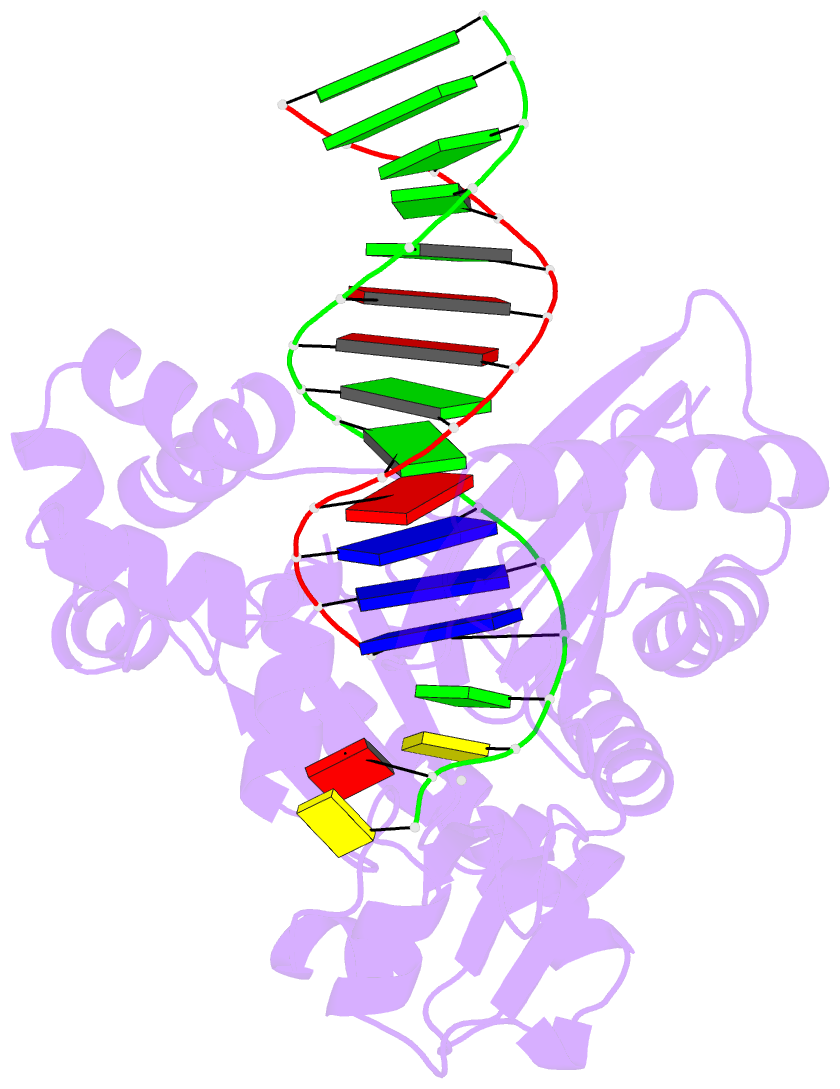
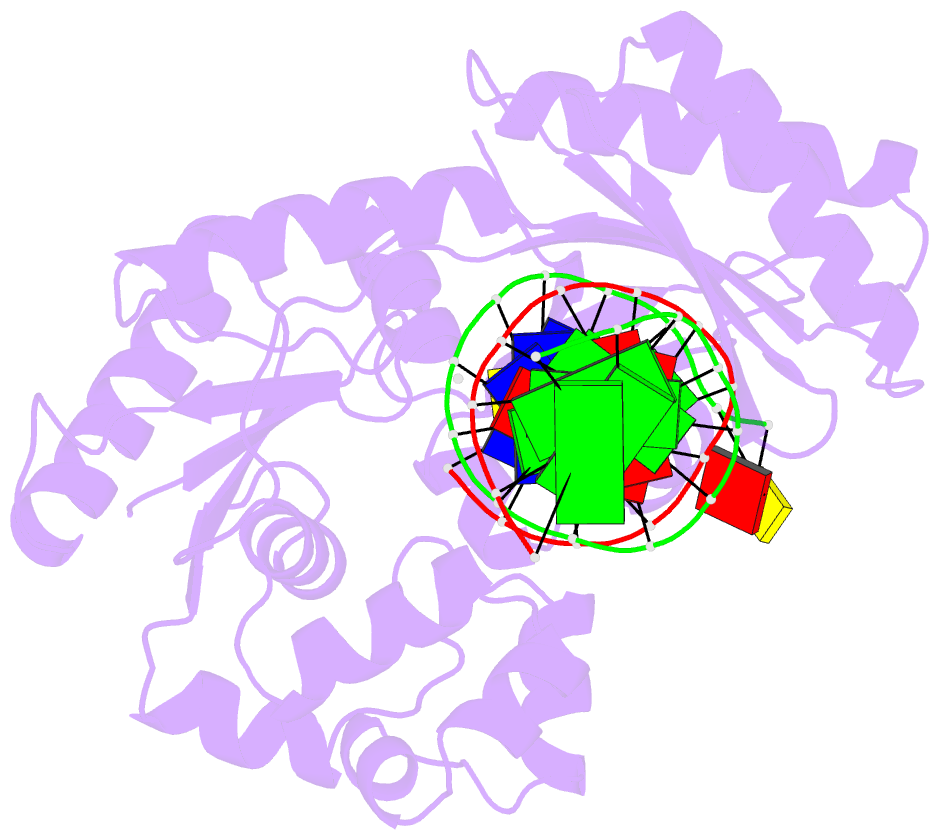
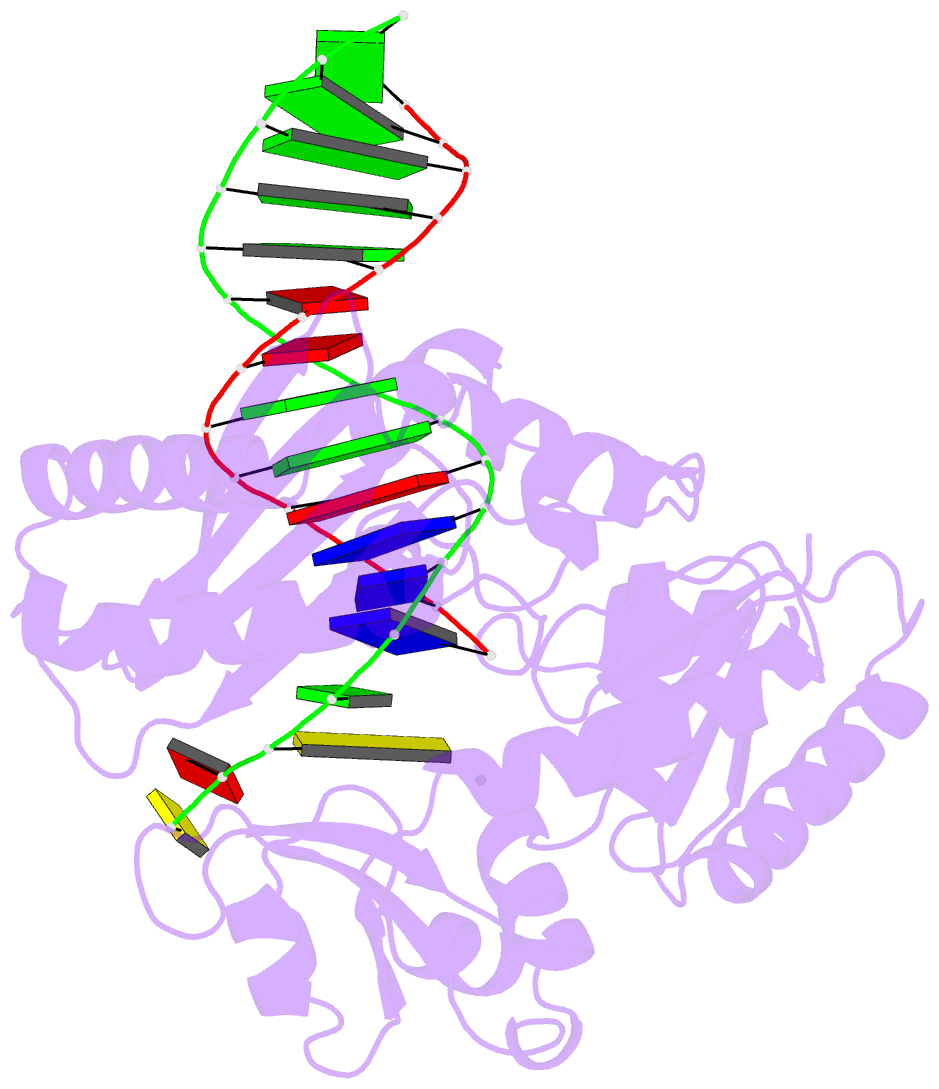
Summary information and primary citation
- PDB-id
- 2r8g; SNAP-derived features in text and JSON formats;
DNAproDB
- Class
- replication, transferase-DNA
- Method
- X-ray (2.7 Å)
- Summary
- Selectivity of nucleoside triphosphate incorporation opposite 1,n2-propanodeoxyguanosine (pdg) by the sulfolobus solfataricus DNA polymerase dpo4 polymerase
- Reference
- Wang Y, Musser SK, Saleh S, Marnett LJ, Egli M, Stone MP (2008): "Insertion of dNTPs opposite the 1,N2-propanodeoxyguanosine adduct by Sulfolobus solfataricus P2 DNA polymerase IV." Biochemistry, 47, 7322-7334. doi: 10.1021/bi800152j.
- Abstract
- 1, N (2)-Propanodeoxyguanosine (PdG) is a stable structural analogue for the 3-(2'-deoxy-beta- d- erythro-pentofuranosyl)pyrimido[1,2-alpha]purin-10(3 H)-one (M 1dG) adduct derived from exposure of DNA to base propenals and to malondialdehyde. The structures of ternary polymerase-DNA-dNTP complexes for three template-primer DNA sequences were determined, with the Y-family Sulfolobus solfataricus DNA polymerase IV (Dpo4), at resolutions between 2.4 and 2.7 A. Three template 18-mer-primer 13-mer sequences, 5'-d(TCACXAAATCCTTCCCCC)-3'.5'-d(GGGGGAAGGATTT)-3' (template I), 5'-d(TCACXGAATCCTTCCCCC)-3'.5'-d(GGGGGAAGGATTC)-3' (template II), and 5'-d(TCATXGAATCCTTCCCCC)-3'.5'-d(GGGGGAAGGATTC)-3' (template III), where X is PdG, were analyzed. With templates I and II, diffracting ternary complexes including dGTP were obtained. The dGTP did not pair with PdG, but instead with the 5'-neighboring template dC, utilizing Watson-Crick geometry. Replication bypass experiments with the template-primer 5'-TCACXAAATCCTTACGAGCATCGCCCCC-3'.5'-GGGGGCGATGCTCGTAAGGATTT-3', where X is PdG, which includes PdG in the 5'-CXA-3' template sequence as in template I, showed that the Dpo4 polymerase inserted dGTP and dATP when challenged by the PdG adduct. For template III, in which the template sequence was 5'-TXG-3', a diffracting ternary complex including dATP was obtained. The dATP did not pair with PdG, but instead with the 5'-neighboring T, utilizing Watson-Crick geometry. Thus, all three ternary complexes were of the "type II" structure described for ternary complexes with native DNA [Ling, H., Boudsocq, F., Woodgate, R., and Yang, W. (2001) Cell 107, 91-102]. The PdG adduct remained in the anti conformation about the glycosyl bond in each of these threee ternary complexes. These results provide insight into how -1 frameshift mutations might be generated for the PdG adduct, a structural model for the exocylic M 1dG adduct formed by malondialdehyde.